太陽光不僅是植物生長的能量來源,還是調(diào)節(jié)植物生長發(fā)育過程的關(guān)鍵信號。光敏色素是植物的紅光/遠(yuǎn)紅光感受器,作為植物的“眼睛”扮演重要角色。當(dāng)下全球氣候變化愈加頻繁、耕地資源逐步縮緊,維護(hù)全球糧食安全是一嚴(yán)重挑戰(zhàn)。過去近一個世紀(jì),農(nóng)作物產(chǎn)量的不斷提高得益于越來越耐密植的作物新品種。將來作物產(chǎn)量進(jìn)一步提高也將繼續(xù)依賴于培育更耐密集種植的新品種。由于密植遮蔭環(huán)境中植物的感光機(jī)理和耐密高產(chǎn)性狀均和植物光敏色素蛋白息息相關(guān),闡明植物光敏色素響應(yīng)以及傳遞光信號的機(jī)制將有助于改善作物密植性能及對復(fù)雜環(huán)境的適應(yīng)性,從而為糧食安全的保障做出貢獻(xiàn)。
高等植物主要編碼以光敏色素A(phyA)和光敏色素B(phyB)為代表的兩類光敏色素,其中phyB是介導(dǎo)可逆紅光響應(yīng)的主要紅光受體。光敏色素通過發(fā)色團(tuán)PΦB在紅光吸收態(tài)(Pr,基態(tài))以及遠(yuǎn)紅光吸收態(tài)(Pfr,激活態(tài))之間進(jìn)行可逆轉(zhuǎn)變。擬南芥phyB被光激活后,可以直接與一類光敏色素互作因子(phytochrome-interacting factor,PIF)互作,傳遞光信號并調(diào)控下游基因表達(dá),促進(jìn)光形態(tài)建成。因此phyB和PIFs構(gòu)成了植物響應(yīng)周圍光環(huán)境的關(guān)鍵信號模塊。擬南芥中8個PIF成員(PIF1-8)均包含兩個重要結(jié)構(gòu)域:N端為結(jié)合phyB-Pfr的激活態(tài)結(jié)合域(Active-PHYB Binding motif,APB),其C端為結(jié)合DNA的bHLH二聚化結(jié)構(gòu)域。在黑暗條件下,phyB-Pr定位于細(xì)胞質(zhì)中,PIF1/3/4/5在核中作用驅(qū)動幼苗暗形態(tài)建成的發(fā)育過程(下胚軸伸長,子葉閉合)。一旦幼苗感知紅光,phyB-Pfr入核進(jìn)而負(fù)調(diào)控上述PIFs,抑制下胚軸伸長和促進(jìn)子葉展開,并維持植物光生長形態(tài)。盡管2022年美國Vierstra團(tuán)隊在Nature發(fā)文揭示了phyB-Pr的結(jié)構(gòu),但phyB-Pfr及其識別PIF的結(jié)構(gòu)生物學(xué)基礎(chǔ)及其調(diào)控機(jī)理仍不清楚。
2024年9月24日,北京大學(xué)現(xiàn)代農(nóng)學(xué)院王繼縱課題組與鄧興旺課題組合作在Cell發(fā)表了題為Light-induced remodeling of phytochrome B enables signal transduction by phytochrome-interacting factor的研究論文,揭示了長期以來期待的phyB光信號轉(zhuǎn)導(dǎo)的最初反應(yīng)機(jī)制。

該研究解析了模式植物擬南芥光激活態(tài)phyB-Pfr以及不依賴于光的組成型激活突變體phyBY276H分別結(jié)合下游信號分子PIF6的復(fù)合物高分辨率冷凍電鏡結(jié)構(gòu)(分辨率依次為3.1 ?,3.2 ?)。基于分子結(jié)構(gòu)分析、結(jié)合突變體蛋白光譜性質(zhì)測定、體外以及半體外生化實驗表征、以及轉(zhuǎn)基因植物的表型分析,揭示了光激活phyB由Pr轉(zhuǎn)變?yōu)镻fr的詳細(xì)分子機(jī)制,并提出了phyB和PIF之間“誘導(dǎo)-契合”的相互作用模型,填補(bǔ)了植物光敏色素信號傳導(dǎo)機(jī)制研究的一個關(guān)鍵空白。
由于全長PIFs蛋白存在大量無序結(jié)構(gòu),體外重組表達(dá)以及組裝phyB-PIF全長蛋白復(fù)合物存在相當(dāng)大的困難。考慮到系列文獻(xiàn)報道中PIFs的APB motif(N端100個氨基酸)足以介導(dǎo)其與phyB在體內(nèi)及體外互作,研究者選取了結(jié)合能力最強(qiáng)的PIF6-APB(PIF6-100,僅N端100個氨基酸)進(jìn)行phyB-PIF的復(fù)合物組裝并制備冷凍樣品。最終成功解析得到了phyB-Pfr-PIF6以及缺失HKRD結(jié)構(gòu)域(僅含N端908個氨基酸)的phyBY276H-908-PIF6復(fù)合物的高分辨率冷凍電鏡結(jié)構(gòu)。結(jié)構(gòu)分析表明phyB-Pfr在光激活之后發(fā)生大規(guī)模結(jié)構(gòu)重排,其光感應(yīng)模塊(photosensory module, PSM)從Pr狀態(tài)下“頭對尾”轉(zhuǎn)變?yōu)?/span>Pfr狀態(tài)下“頭對頭”二聚體,且PIF6-APB單體結(jié)合在phyB-Pfr二聚體界面的一側(cè),組成phyB-PIF6三聚體。此外,phyBY276H-908-PIF6復(fù)合物結(jié)構(gòu)與phyB-Pfr-PIF6幾乎完全一致(圖1)。
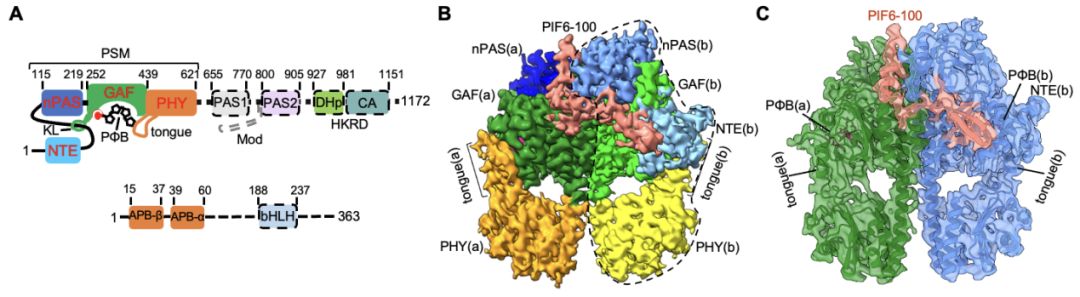
圖1. phyB-Pfr-PIF6(B)和phyBY276H-PIF6(C)均為不對稱三聚體
為了闡明光激活驅(qū)動phyB的結(jié)構(gòu)重排,作者細(xì)致比對了之前文獻(xiàn)報道的phyB-Pr及本研究獲得的phyB-Pfr結(jié)構(gòu),發(fā)現(xiàn)發(fā)色團(tuán)PΦB分子在吸收紅光后,其D環(huán)翻轉(zhuǎn)了180?,并且和口袋中的系列氨基酸重新建立互作網(wǎng)絡(luò)(圖2A,左),最終導(dǎo)致PHY結(jié)構(gòu)域里與其互作的舌狀突出結(jié)構(gòu)發(fā)生由β片層到α螺旋的構(gòu)象轉(zhuǎn)變(圖2A,右)。結(jié)構(gòu)分析表明,絲氨酸S584對于穩(wěn)定α螺旋形式的舌狀結(jié)構(gòu)非常重要(圖2A,右),進(jìn)一步的體外生化以及轉(zhuǎn)基因植物表型分析證明了S584對于維系phyB-Pfr激活狀態(tài)和phyB信號通路至關(guān)重要(圖2B)。
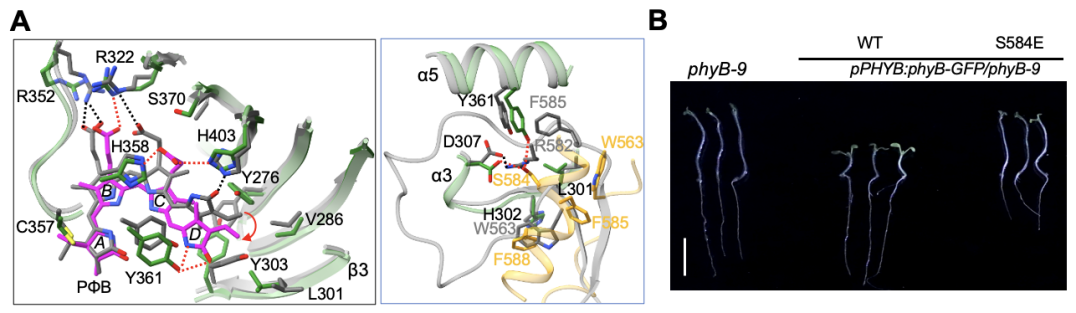
圖2. 光誘導(dǎo)PΦB分子(A,左)和PHY結(jié)構(gòu)域舌狀突出結(jié)構(gòu)(A,右)的構(gòu)象轉(zhuǎn)變及功能驗證(B)
在Pr狀態(tài)下,phyB的C末端結(jié)構(gòu)域PAS2以及HKRD與N末端結(jié)構(gòu)域GAF和PHY互作以維系“頭對尾”二聚體構(gòu)象。而在phyB-Pfr中,其PHY結(jié)構(gòu)域的舌狀突出結(jié)構(gòu)轉(zhuǎn)變?yōu)?/span>α螺旋后,會直接和Pr狀態(tài)里的PAS2結(jié)構(gòu)域發(fā)生空間沖突(圖3A,左;圖3B,左),從而破壞Pr狀態(tài)下PAS2和PHY結(jié)構(gòu)域之間的廣泛互作,這會進(jìn)一步破壞HKRD和PHY結(jié)構(gòu)域(圖3A,左;圖3B,中),以及HKRD和GAF結(jié)構(gòu)域(圖3A,右;圖3B,右)之間的分子內(nèi)互作,這一系列的構(gòu)象變化完全打破Pr狀態(tài)下的“頭對尾”結(jié)構(gòu),最終激活phyB。此外,缺失HKRD結(jié)構(gòu)域的phyB截短體蛋白(phyB-908,僅含N端908個氨基酸)在光激活之后傾向于單體形式存在(圖3C),進(jìn)一步佐證了phyB-Pr在光激活后,原來由N端和C端模塊互作維系的“頭對尾”二聚體被打破,進(jìn)而形成phyB-Pfr。
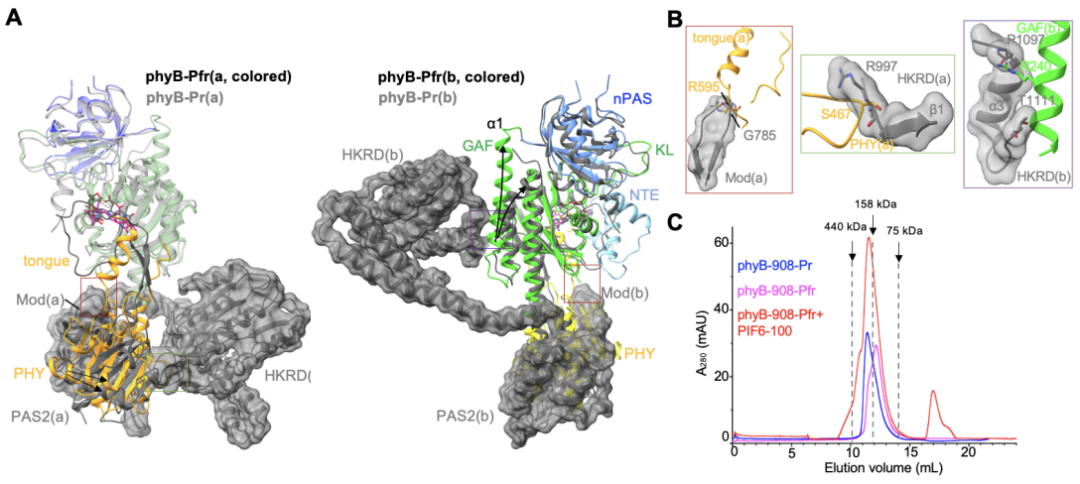
圖3. PHY 結(jié)構(gòu)域舌狀突出結(jié)構(gòu)的構(gòu)象轉(zhuǎn)變驅(qū)動phyB-Pr重構(gòu)形成phyB-Pfr(A,B)及生化驗證(C)
光激活后的phyB-Pfr分別利用其N末端的NTE結(jié)構(gòu)域以及由整體N末端結(jié)構(gòu)域形成的“頭對頭”二聚體,來識別和結(jié)合PIF6-APB的N末端和C末端(圖4A)。PIF6-APB的N末端通過形成一個β發(fā)夾結(jié)構(gòu)(PIF6-APB-β)與phyB-Pfr的NTE結(jié)構(gòu)域發(fā)生廣泛互作(圖4B),而phyB-Pfr二聚體結(jié)構(gòu)中另一分子phyB并不具備穩(wěn)定結(jié)構(gòu)的NTE,表明NTE的穩(wěn)定結(jié)構(gòu)是由PIF6-APB的結(jié)合所誘導(dǎo)形成的。此外,PIF6-APB的C末端通過形成一個α螺旋結(jié)構(gòu)(PIF6-APB-α)同時與兩個phyB-Pfr的N末端結(jié)構(gòu)域產(chǎn)生廣泛互作(圖4C),印證了PIF6-100促進(jìn)的phyB-908-Pfr的二聚化(圖3C)。作者進(jìn)一步通過系列生化、光譜實驗表征、結(jié)合轉(zhuǎn)基因植物的表型明確了NTE結(jié)構(gòu)域以及N末端結(jié)構(gòu)域介導(dǎo)的phyB-Pfr二聚化對于phyB-PIF互作的重要性。
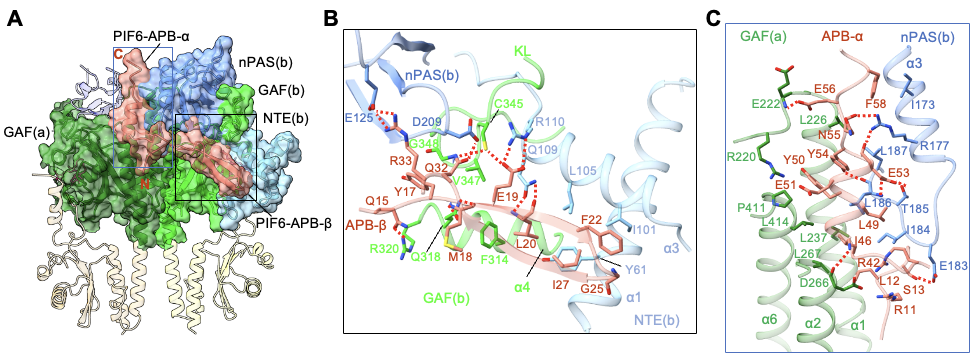
圖4. phyB-Pfr的N末端結(jié)構(gòu)域?qū)?/span>PIF-APB的特異性識別(A)以及NTE結(jié)構(gòu)域特異性識別PIF-APB的N末端β發(fā)夾結(jié)構(gòu)(B),phyB-Pfr二聚體特異性識別PIF-APB的C末端α螺旋結(jié)構(gòu)(C)
兩個phyB-Pfr的結(jié)構(gòu)比對表明phyB-PIF6復(fù)合物結(jié)構(gòu)中的phyB-Pfr形成了不對稱的二聚體(圖5A),這些不對稱性導(dǎo)致phyB-Pfr二聚體的另一側(cè)界面沒有足夠空間與PIF6-APB-β以及PIF6-APB-α形成合適的互作(圖5B)。作者進(jìn)一步通過pull-down以及Co-IP assay評估了溶液狀態(tài)下phyB和PIF-APB以及PIF-FL的互作模式,生化數(shù)據(jù)表明PIF3和PIF6的APB motif以及PIF1和PIF3的全長蛋白均可以和phyB-Pfr二聚體形成摩爾比為1:2(2:4)的復(fù)合物。
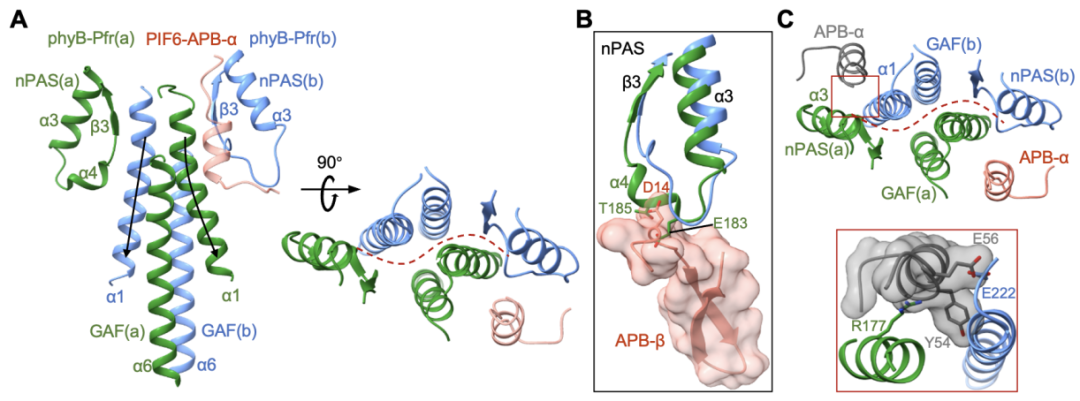
圖5. 不對稱的phyB-Pfr二聚體只能結(jié)合一個PIF6-APB單體
總的來說,這項研究揭示了“頭對頭”激活態(tài)phyB二聚體識別結(jié)合PIF6-APB的復(fù)合物結(jié)構(gòu),解決了光敏色素研究中光如何誘導(dǎo)phyB變構(gòu)激活并轉(zhuǎn)導(dǎo)PIF信號的關(guān)鍵問題(圖6),為作物感光性狀的改造以及phyB相關(guān)光控基因表達(dá)的“光遺傳學(xué)”工具開發(fā)提供了分子水平的精細(xì)圖紙。
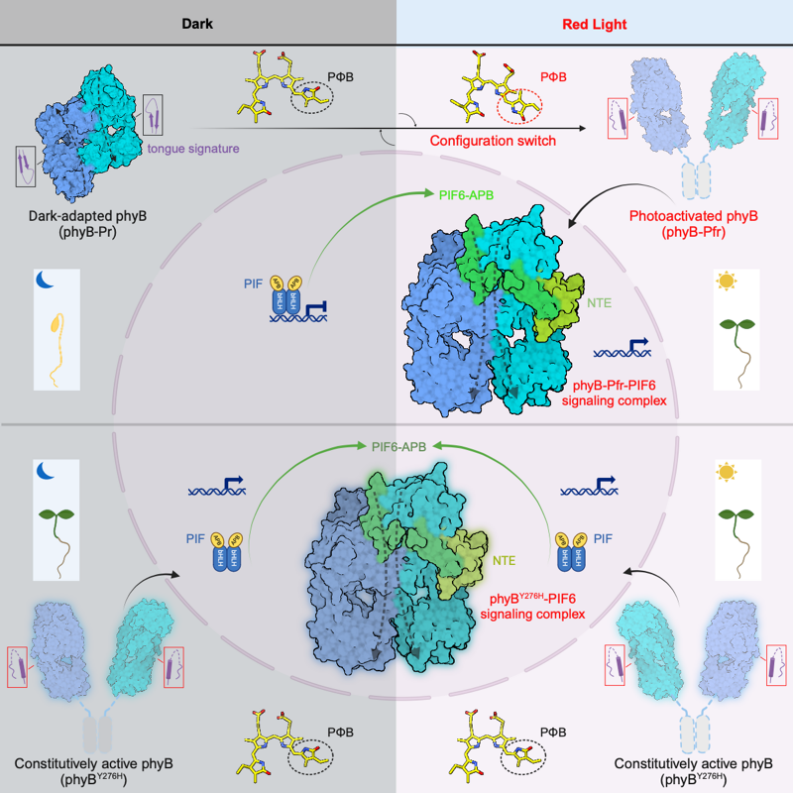
圖6. 紅光激活phyB并轉(zhuǎn)導(dǎo)PIF信號的結(jié)構(gòu)機(jī)制模型
北京大學(xué)現(xiàn)代農(nóng)學(xué)院、北京大學(xué)現(xiàn)代農(nóng)業(yè)研究院王繼縱研究員和鄧興旺教授為該論文的通訊作者。北京大學(xué)前沿交叉學(xué)科研究院博士研究生王征東和宋艷萍、北京大學(xué)現(xiàn)代農(nóng)業(yè)研究院科研助理王文鳳和趙迪迪、以及北京大學(xué)現(xiàn)代農(nóng)業(yè)研究院林曉莉博士為論文共同第一作者。北京大學(xué)現(xiàn)代農(nóng)業(yè)研究院趙珺博士、遲程博士和高級工程師徐斌、中國科學(xué)院遺傳與發(fā)育生物學(xué)研究所博士研究生沈萌對本研究也做出了重要貢獻(xiàn)。冷凍樣品制備、樣品篩選和數(shù)據(jù)收集在研究院生物微觀結(jié)構(gòu)研究平臺完成。該研究得到了國家自然科學(xué)基金、國家重點研發(fā)項目、山東省重點研發(fā)項目、山東省科技創(chuàng)新基金、中科協(xié)青年人才托舉工程、山東省泰山青年學(xué)者項目、北京大學(xué)現(xiàn)代農(nóng)業(yè)研究院、濰坊現(xiàn)代農(nóng)業(yè)山東省實驗室、小麥育種全國重點實驗室、北京大學(xué)蛋白質(zhì)與植物基因研究國家重點實驗室相關(guān)經(jīng)費的資助。
論文鏈接:https://www.cell.com/cell/fulltext/S0092-8674(24)01023-7
專家觀點
萬建民 院士(中國農(nóng)業(yè)科學(xué)院作物科學(xué)研究)
光照是植物生長的能量來源,對農(nóng)作物產(chǎn)量起著決定性作用。在農(nóng)業(yè)生產(chǎn)中,提高作物種植密度是增產(chǎn)的重要途徑。然而,隨著密度增加,作物植株之間相互遮陰會加劇,造成植株莖稈增高、變?nèi)酰装l(fā)生倒伏、小穗和抗逆性減弱等突出問題。因此,闡明植物感知光照的分子機(jī)理對提高作物光適應(yīng)能力和耐密性等具有十分重要的意義。光敏色素是植物的紅光、遠(yuǎn)紅光受體,是影響作物響應(yīng)光照變化的關(guān)鍵功能蛋白。自1960年首次以“光敏色素”命名這類光受體以來,研究人員一直好奇光敏色素感知紅光/遠(yuǎn)紅光信號的機(jī)制。早在上世紀(jì)80年代末,研究人員在植物光敏色素phyA的電鏡照片中就看到了“Y”字形的蛋白顆粒。但是由于光敏色素屬于多結(jié)構(gòu)域光受體蛋白,植物內(nèi)源含量較低,且重組表達(dá)困難,研究人員主要通過對相對容易獲得的微生物來源光敏色素進(jìn)行結(jié)構(gòu)表征,植物光敏色素的結(jié)構(gòu)研究相對停滯。直到2014年,PNAS雜志發(fā)表了基態(tài)phyB光感應(yīng)模塊截短體的晶體結(jié)構(gòu),2022年以來,來自于美國以及中國的多家單位相繼報道了全長phyB及phyA基態(tài)的結(jié)構(gòu),從而逐漸建立了光敏色素基態(tài)結(jié)構(gòu)特性的認(rèn)識。但植物光敏色素激活態(tài)結(jié)構(gòu)及光信號原初轉(zhuǎn)導(dǎo)的機(jī)制仍不清楚,限制了基于激活態(tài)phyB結(jié)構(gòu)的光信號轉(zhuǎn)導(dǎo)機(jī)制研究。
這項研究首次報道了激活態(tài)植物光敏色素phyB結(jié)合標(biāo)志性下游信號分子PIF的復(fù)合物結(jié)構(gòu),揭示了廣泛研究的不依賴與光照的組成型激活突變體phyBY276H結(jié)合PIF的復(fù)合物結(jié)構(gòu),并結(jié)合多種研究方法,闡明了光照引起的激活態(tài)phyB結(jié)構(gòu)重排以及光敏色素蛋白和發(fā)色團(tuán)分子之間相互協(xié)調(diào)的動態(tài)變化機(jī)制。這項研究充分回答了長期以來研究人員關(guān)注的phyB感光激活機(jī)理,填補(bǔ)了激活態(tài)植物光敏色素結(jié)構(gòu)研究的空白,是生物學(xué)基礎(chǔ)研究的重大突破。同時,光敏色素激活機(jī)制的解析為精準(zhǔn)調(diào)控植物光響應(yīng)建立了基礎(chǔ),對定向改良玉米、小麥、水稻等重要作物的感光性狀、培育耐密高產(chǎn)品種意義重大。
王海洋 教授(華南農(nóng)業(yè)大學(xué))
培育耐密植作物是提高作物單產(chǎn)的有效技術(shù)措施。株型是決定作物耐密性的關(guān)鍵因素之一。然而,密植容易誘發(fā)植物產(chǎn)生避蔭反應(yīng),導(dǎo)致植株徒長、莖稈變細(xì)、根系發(fā)育減弱、增加倒伏風(fēng)險等不利后果。作為植物響應(yīng)紅光/遠(yuǎn)紅光信號的主要紅光受體,光敏色素phyB對于植物應(yīng)對外界光環(huán)境變化的適應(yīng)性生長(株高、葉夾角、開花時間等)至關(guān)重要,從而影響作物的株型、光合效率和產(chǎn)量。phyB被紅光激活后,通過與PIF蛋白直接互作傳遞光信號,進(jìn)而啟動和維持光形態(tài)建成的發(fā)育過程。然而自1998年發(fā)現(xiàn)PIF是激活態(tài)光敏色素結(jié)合蛋白以來,雖然鑒定了部分影響phyB-PIF互作的關(guān)鍵位點,但激活態(tài)phyB與PIF蛋白互作的結(jié)構(gòu)基礎(chǔ)和調(diào)控機(jī)理目前仍然未知。這項研究通過解析了模式植物擬南芥光激活態(tài)phyB-Pfr以及不依賴于光的組成型激活突變體phyBY276H分別結(jié)合下游信號分子PIF6的復(fù)合物高分辨率冷凍電鏡結(jié)構(gòu),揭示了phyB基態(tài)(Pr)在光激活后,其光感應(yīng)模塊(PSM)從Pr狀態(tài)下“頭對尾”轉(zhuǎn)變?yōu)?/span>Pfr狀態(tài)下“頭對頭”二聚體。由于phyB-Pfr二聚體的不對稱性,導(dǎo)致其僅一側(cè)能夠結(jié)合PIF并形成phyB-Pfr-PIF的三聚體。作者進(jìn)一步結(jié)合突變體蛋白光譜性質(zhì)測定、體外以及半體外生化實驗表征、以及轉(zhuǎn)基因植物的表型分析,驗證了絲氨酸S584、NTE結(jié)構(gòu)域以及N末端結(jié)構(gòu)域?qū)τ诮閷?dǎo)phyB-Pfr二聚化,phyB-PIF互作和phyB信號通路的重要性。該研究首次揭示了phyB光激活及最初光信號傳遞(與PIF互作)的結(jié)構(gòu)生物學(xué)基礎(chǔ),是近年來植物光敏色素信號傳導(dǎo)研究領(lǐng)域取得的又一個重大突破。研究成果可為培育耐蔭、耐密、抗倒伏、高光效作物新品種提供重要理論指導(dǎo)和基因資源。